Life in the Soil
A healthy soil food web supports a consistent release of minerals and nutrients into the soil to support plant growth. In a healthy soil the minerals and nutrients in a soil are locked up in large soil aggregates, these can be broken down by bacteria and fungi in the soil to provide a slow release fertiliser for plant growth. In exchange, plants provide the bacteria and fungi with sugars produced by photosynthesis through their roots. This creates a dynamic and complex food web in the soil which sustains plant growth without any chemical fertilisers.
Soil contains a complex ecosystem of microorganisms that until recently has been little studied. It is often said that a handful of soil has more living organisms than there are people on planet Earth. When the biology in soil is balanced, soil can produce healthy plants season after season. These naturally occurring organisms convert mineral and organic material into nutrients that plants require, creating a symbiotic relationship between plant and soil.
Soil organic matter (SOM) is defined as a complex mixture of plant (flora) and animal (fauna) residues at various stages of decomposition, root exudates and soil biota. Soil biota comprises all the organisms living within the soil ranging from soil microbial fungi and bacteria to protozoa and nematodes. Together they are responsible for the decomposition of organic materials and cycling of nutrients within the soil.
Soil organisms participate in the genesis of the habitat wherein they live. They, together with the total biota and especially the higher vegetation, constitute one of the five interactive factors of soil formation; the other four are climate, topography, parent material and time.
The physical and chemical breakdown of rocks to fine particles with large surface areas and the accompanying release of plant nutrients initiate the soil-forming process.
The initial colonisers are usually organisms capable both of photosynthesis and nitrogen fixation. These are predominantly cyanobacteria, also known as the blue-green algae. Lichens in which the algal partner is a cyanobacterium have also an important role at these early stages of soil development.
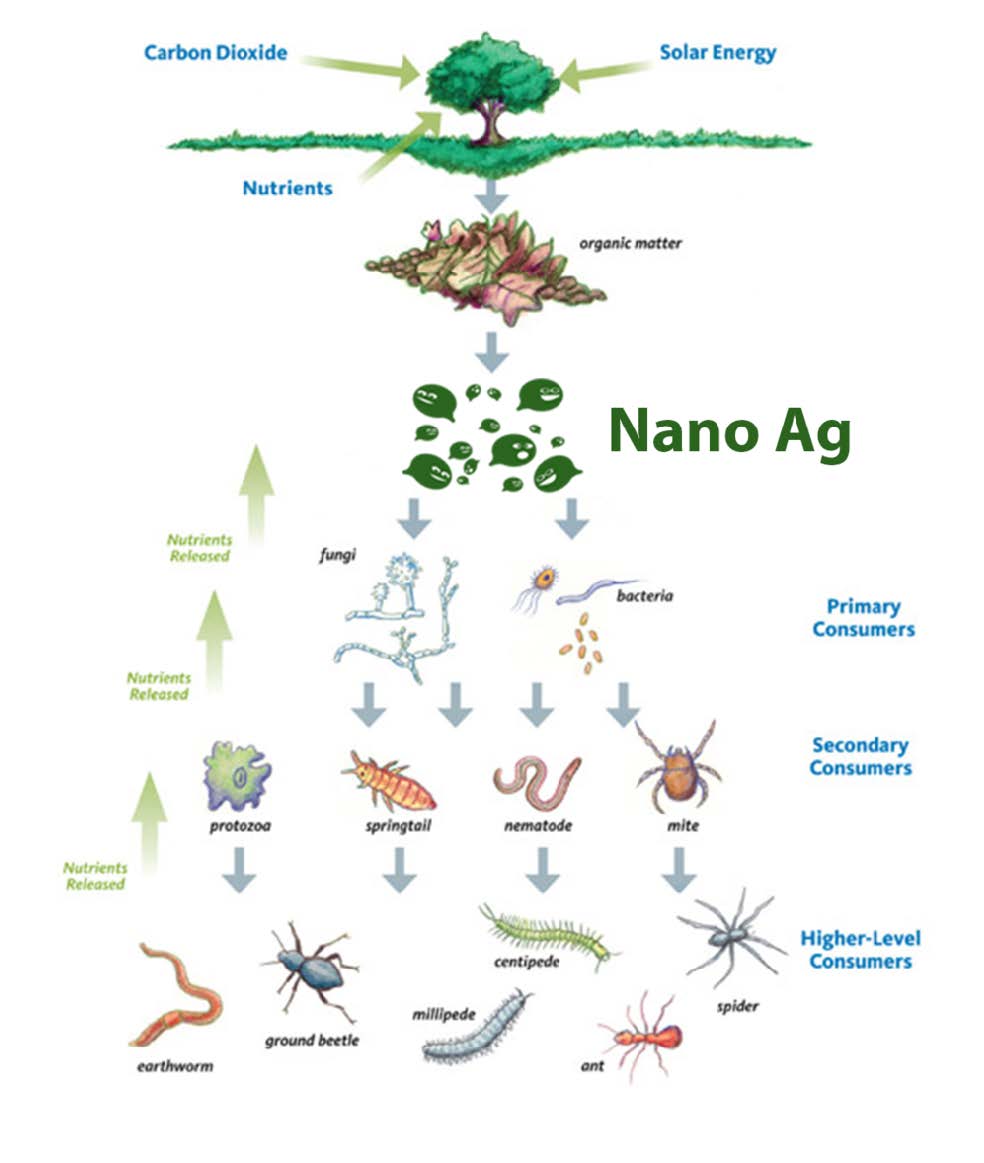
Life in the soil can be organised into trophic levels. At the base of the trophic levels lies the soil microbial population which degrades plant, animal and microbial bodies, and also serves as the food source for some of the levels above it. For example, soil protozoa consume enormous numbers of bacteria and even some fungal spores.These in turn are consumed by still larger soil animals (nematodes, mites, etc.) which in turn are eaten by still larger animals (e.g. worms and insects). Thus, nutrients flow through this microbial food web which lies at the heart of controlling soil fertility and plant productivity in the absence of external inputs such as fertilizers.
Microscopic life forms in the soil are generally called the "soil microflora" (though strictly speaking, not all are plants in the true sense of the word) and the larger animals are called macrofauna. Soil animals, especially, the earthworms and some insects tend to affect the soil favorably through their burrowing and feeding activities which tend to improve aeration and drainage through structural modifications of the soil.
Plant or animal material and faeces fall to the soil and may be attacked first by fungi or bacteria which rapidly decompose the carbohydrates, proteins and fats. Alternatively litter may be eaten by soil animals. This breaks it down into smaller pieces more readily attacked by micro-organisms and distributed more widely in the soil. The fungi that grow on the more resistant lignin material may themselves be eaten by animals such as mites, springtails and nematodes, which, when they die, are again food for further bacteria. These animals are the primary colonisers of cellulose and may be eaten by protozoa, which in turn are eaten by their predators or die providing substrates for bacteria and fungi. Thus, decomposition processes in soils involve the total number of soil organisms in a complex network of food-chains which eventually return the more complex organic components to their simple inorganic constituents.
There are complex interaction between soil organisms. Many of these interactions are based around symbiotic relationships where each organism's benefits the other. The most widely recognized example is that between nitrogen fixing Rhizobium bacteria that live in root nodules of legume host plants and exchange with those plants nitrates created from atmospheric nitrogen in exchange for sugars produced by the host plant. Nearly two-thirds of the world's nitrogen supply is from biological nitrogen fixation. Legumes have been used since the beginning of recorded history as "soil improving" crops known as "green manures". Green manuring is the practice of growing a legume species for the sole purpose of returning it to the soil to serve as a source of nitrogen for an ensuing crop.
Soil microorganisms occur in huge numbers and display an enormous diversity of forms and functions. Major microbial groups in soil are bacteria, fungi, algae (including cyanobacteria) and protozoa. Soil microbes can occur in numbers ranging up to several million or more in a gram of fertile soil (a volume approximately that of a red kidney bean). Bacteria are the most numerous of the soil microbes. However, It is the soil fungi which contribute the most biomass among the microbial groups. A fungus, Armillaria bulbosa, discovered in the U.S. could turn out to be earth's largest living creature on Earth. The microscopic, branched filaments (called hyphae) of the fungus occupy a 14.8 ha (37-acre) area of land. Careful genetic analysis has shown the filaments constitute a single organism. Scientists estimate that the portion of the fungus they have been able to identify may weigh as much as 100 tons, slightly less than a blue whale. Imagine the biochemical capacity of a soil microorganism this large!
Soil contains a complex ecosystem of microorganisms that until recently has been little studied. It is often said that a handful of soil has more living organisms than there are people on planet Earth. When the biology in soil is balanced, soil can produce healthy plants season after season. These naturally occurring organisms convert mineral and organic material into nutrients that plants require, creating a symbiotic relationship between plant and soil.
Soil organic matter (SOM) is defined as a complex mixture of plant (flora) and animal (fauna) residues at various stages of decomposition, root exudates and soil biota. Soil biota comprises all the organisms living within the soil ranging from soil microbial fungi and bacteria to protozoa and nematodes. Together they are responsible for the decomposition of organic materials and cycling of nutrients within the soil.
Soil organisms participate in the genesis of the habitat wherein they live. They, together with the total biota and especially the higher vegetation, constitute one of the five interactive factors of soil formation; the other four are climate, topography, parent material and time.
The physical and chemical breakdown of rocks to fine particles with large surface areas and the accompanying release of plant nutrients initiate the soil-forming process.
The initial colonisers are usually organisms capable both of photosynthesis and nitrogen fixation. These are predominantly cyanobacteria, also known as the blue-green algae. Lichens in which the algal partner is a cyanobacterium have also an important role at these early stages of soil development.
Life in the soil can be organised into trophic levels. At the base of the trophic levels lies the soil microbial population which degrades plant, animal and microbial bodies, and also serves as the food source for some of the levels above it. For example, soil protozoa consume enormous numbers of bacteria and even some fungal spores.These in turn are consumed by still larger soil animals (nematodes, mites, etc.) which in turn are eaten by still larger animals (e.g. worms and insects). Thus, nutrients flow through this microbial food web which lies at the heart of controlling soil fertility and plant productivity in the absence of external inputs such as fertilizers.
Microscopic life forms in the soil are generally called the "soil microflora" (though strictly speaking, not all are plants in the true sense of the word) and the larger animals are called macrofauna. Soil animals, especially, the earthworms and some insects tend to affect the soil favorably through their burrowing and feeding activities which tend to improve aeration and drainage through structural modifications of the soil.
Plant or animal material and faeces fall to the soil and may be attacked first by fungi or bacteria which rapidly decompose the carbohydrates, proteins and fats. Alternatively litter may be eaten by soil animals. This breaks it down into smaller pieces more readily attacked by micro-organisms and distributed more widely in the soil. The fungi that grow on the more resistant lignin material may themselves be eaten by animals such as mites, springtails and nematodes, which, when they die, are again food for further bacteria. These animals are the primary colonisers of cellulose and may be eaten by protozoa, which in turn are eaten by their predators or die providing substrates for bacteria and fungi. Thus, decomposition processes in soils involve the total number of soil organisms in a complex network of food-chains which eventually return the more complex organic components to their simple inorganic constituents.
There are complex interaction between soil organisms. Many of these interactions are based around symbiotic relationships where each organism's benefits the other. The most widely recognized example is that between nitrogen fixing Rhizobium bacteria that live in root nodules of legume host plants and exchange with those plants nitrates created from atmospheric nitrogen in exchange for sugars produced by the host plant. Nearly two-thirds of the world's nitrogen supply is from biological nitrogen fixation. Legumes have been used since the beginning of recorded history as "soil improving" crops known as "green manures". Green manuring is the practice of growing a legume species for the sole purpose of returning it to the soil to serve as a source of nitrogen for an ensuing crop.
Soil microorganisms occur in huge numbers and display an enormous diversity of forms and functions. Major microbial groups in soil are bacteria, fungi, algae (including cyanobacteria) and protozoa. Soil microbes can occur in numbers ranging up to several million or more in a gram of fertile soil (a volume approximately that of a red kidney bean). Bacteria are the most numerous of the soil microbes. However, It is the soil fungi which contribute the most biomass among the microbial groups. A fungus, Armillaria bulbosa, discovered in the U.S. could turn out to be earth's largest living creature on Earth. The microscopic, branched filaments (called hyphae) of the fungus occupy a 14.8 ha (37-acre) area of land. Careful genetic analysis has shown the filaments constitute a single organism. Scientists estimate that the portion of the fungus they have been able to identify may weigh as much as 100 tons, slightly less than a blue whale. Imagine the biochemical capacity of a soil microorganism this large!
Video Resources
There are a lot of excellent videos on this topic and new discoveries coming out all the time. Working with successional plant communities to build healthy soil and create more stable and diverse perennial food producing systems is really a central concept of permacultre design.
|
|
|
|
|
|
|
|
|
Soil Food Web
Pores as Biota Habita
Soil pores are an important component of soils. Mineral soils usually consist of 45-55 % (on volume basis) of pores of different sizes.
Soil biota is largely influenced by soil-water relationships, as water in soil affects soil aeration and, therefore, the types of soil microorganisms present in soil, their population size and activities. According to capillary laws, only pores having diameters smaller than a critical value are filled with water at a given water potential.
Soil atmosphere is strongly influenced by respiration of plant roots and soil organisms.
Waterlogging conditions are frequently found in poorly-drained, fine textured soils, but they can also occur in soils normally well drained. Poorly-aerated soils are inhibitory to O2-dependent plant and microbial respiration; however, anaerobic microorganisms grow under such conditions.
Microorganisms occupy less than 1% of the total available pore space. Pore neck sizes determine accessibility to pores by organisms according to their body sizes.
Different groups of microorganisms are found at different depths, as a function of their nutritional requirements and tolerances.
The soil region under immediate influence of plant roots (rhizosphere) there are very high microbial populations and these are distinct from the rest of soil.
Soil biota is largely influenced by soil-water relationships, as water in soil affects soil aeration and, therefore, the types of soil microorganisms present in soil, their population size and activities. According to capillary laws, only pores having diameters smaller than a critical value are filled with water at a given water potential.
Soil atmosphere is strongly influenced by respiration of plant roots and soil organisms.
- A well-aerated soil is one in which gas exchange between the soil air and the atmosphere is sufficiently rapid to prevent a deficiency of O2 or a toxicity of CO2 and thereby permits normal functioning of plant roots and of aerobic microorganisms
- O2 in well-aerated soil is usually above 18-20%; root activity is inhibited when O2 content is below 10%
- CO2 in a dry soil is usually no higher than 1-2% (in the atmosphere is 0.03%); in diffusion-limited systems (waterlogged) it may reach 10%.
Waterlogging conditions are frequently found in poorly-drained, fine textured soils, but they can also occur in soils normally well drained. Poorly-aerated soils are inhibitory to O2-dependent plant and microbial respiration; however, anaerobic microorganisms grow under such conditions.
Microorganisms occupy less than 1% of the total available pore space. Pore neck sizes determine accessibility to pores by organisms according to their body sizes.
Different groups of microorganisms are found at different depths, as a function of their nutritional requirements and tolerances.
The soil region under immediate influence of plant roots (rhizosphere) there are very high microbial populations and these are distinct from the rest of soil.
Bacteria
Bacteria are 10 to 1000 times greater in the rhizosphere, while fungi are not that common, except for symbionts. As plant senesces, pathogens (many fungi) appear in greater number. Protozoa are abundant in the rhizosphere.
The distribution of microbial numbers within a soil profile is highly influenced by the distribution of soil organic matter, which decreases with depth. Other factors that have a role are (i) access to light (this will affect algae numbers; algae needs light and thus is not found at depth), and (ii) presence of a water table (this will affect the dominance of anaerobic microorganisms vs. aerobic microorganisms). Protozoa feed on bacteria and organic matter, so their numbers will also vary accordingly.
Most soil organic matter is derived from the decomposition of plant tissue. Soil organic matter is formed as the plant tissue is incorporated into soil and decomposed by soil organisms colonising it, and then using it as a source of food, amino acids and nutrients for growth. Most plant tissue components are of sufficiently large molecular weight that they are unable to pass directly through the cell wall to be oxidised or hydrolysed directly by intracellular enzyme systems. The decomposer must secrete extracellular hydrolysing or depolymerising enzymes which operate in the soil water film outside their cell walls. These enzymes reduce complex organic molecules to simpler small molecules that can be transported through cell walls. The simpler molecules can then be used as energy sources or as intermediates in the metabolism or synthesis of new biomass.
The decomposition process involves colonisation and growth of decomposer organisms on an organic substrate. Sufficient moisture must be available to cover the decomposer and the substrate with a thin water film. This film provides the transport medium for the organism to invade the substrate, the digestive medium for the extracellular enzymes to work in and the transport medium for the products of decomposition to move to the decomposer and other waste products to leave the site of decomposition (e.g., CO2).
There is a succession of opportunistic decomposer organisms, with new microbial biomass will growing until its supply of substrate is exhausted. As a result of bacterial growth, the population of protozoan graziers will increase. Other predators of the initial decomposers will also increase. Fungi are commonly initial colonisers because their hyphal growth habit allows them to ramify (like a root growing in soil) through the organic substrate whereas bacteria must move where the thin water film allows them to swim. Various types of bacteria and fungi will predate upon each other, each possessing extracellular enzymes or antibiotic agents capable of lysing (breaking down) the cell walls of their prey. The isolation and culture of such organisms on synthetic media has allowed the production of antibiotics (e.g., penicillin and streptomycin) for treatment of human bacterial and fungal infections.
Heterotrophic bacteria
Those which require organic compounds to supply their energy needs are the most common, obtaining these components by secreting externally released extracellular enzymes which attack live and dead animal and plant material. The extracellular enzymes break down the more complex organic materials to simple compounds that can be absorbed by the bacteria.
Fortunately bacteria can attack a wide range of organic substrates, including many organic pesticides and herbicides which are added to crops. Thus toxic substances are prevented from accumulating in the soil.
Bacteria have to compete for inorganic nutrients with plants and fungi and absorb their nitrate, sulphate, phosphate, calcium, potassium, magnesium, and sodium from soil solution.
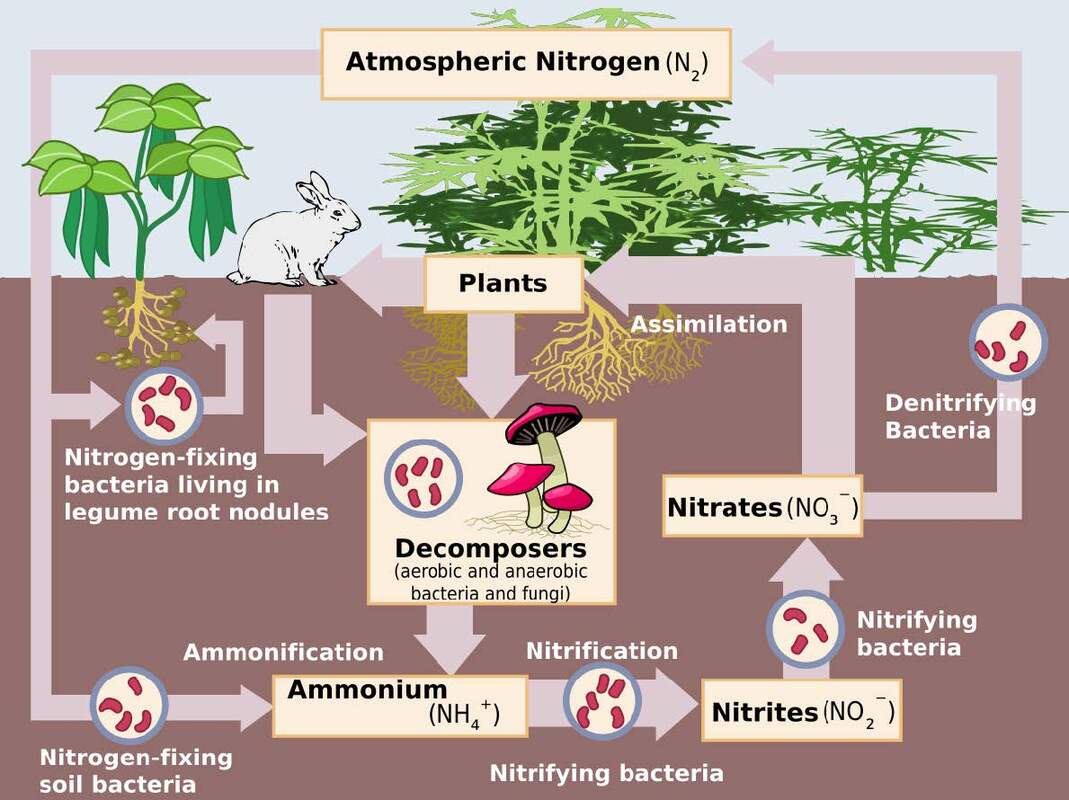
Nitrification.
A process whereby ammonium N is converted to nitrite by the bacteria Nitrosomonas and then nitrate by the bacteria Nitrobacter. Although they grow slowly, large quantities of ammonium can be converted to nitrate in soil. Rates of nitrification are higher in moist neutral to slightly alkaline soils. Oxygen is required for growth. The nitrate produced can then be taken up by plants, lost by leaching or denitrification (an anaerobic process in water logged soil or in the anaerobic centre of soil aggregates in aerobic conditions.
Nitrogen is an essential element for the growth and functioning of all organisms. Soil parent materials contain little or no nitrogen and the major source of nitrogen is inert N2 gas in the atmosphere. This inert N source must initially be converted into a form which can be used by producer organisms (NH3, NH4+ or NO3-). In the natural world this task falls to a few lightning bolts and specialised micro-organisms capable of fixing molecular N2 into NH3. The rest of the organisms on earth are totally dependent upon these micro-organisms and the few producer organisms capable of catalysing the synthesis of essential amino acids. The importance of organisms capable of fixing N2 gas and knowledge of their maintenance are an essential part of global sustainability. In particular the economies of Australia and New Zealand are heavily dependent upon the successful cultivation of legume based pastures.
It has been estimated that in New Zealand approximately 1 million tonnes of N is fixed annually in legume based pastures. In soils where the concentration of inorganic N is high, clovers will use soil N, with the result that there is less biological fixation of atmospheric N. Nitrogen fertiliser addition to pasture soils has often been shown to decrease BNF (Table 4.5.3). With increasing N addition N fixation by clover continue to decrease and the percent decrease varied between 30 to 70% depending on the time of application and the grazing management.
The distribution of microbial numbers within a soil profile is highly influenced by the distribution of soil organic matter, which decreases with depth. Other factors that have a role are (i) access to light (this will affect algae numbers; algae needs light and thus is not found at depth), and (ii) presence of a water table (this will affect the dominance of anaerobic microorganisms vs. aerobic microorganisms). Protozoa feed on bacteria and organic matter, so their numbers will also vary accordingly.
Most soil organic matter is derived from the decomposition of plant tissue. Soil organic matter is formed as the plant tissue is incorporated into soil and decomposed by soil organisms colonising it, and then using it as a source of food, amino acids and nutrients for growth. Most plant tissue components are of sufficiently large molecular weight that they are unable to pass directly through the cell wall to be oxidised or hydrolysed directly by intracellular enzyme systems. The decomposer must secrete extracellular hydrolysing or depolymerising enzymes which operate in the soil water film outside their cell walls. These enzymes reduce complex organic molecules to simpler small molecules that can be transported through cell walls. The simpler molecules can then be used as energy sources or as intermediates in the metabolism or synthesis of new biomass.
The decomposition process involves colonisation and growth of decomposer organisms on an organic substrate. Sufficient moisture must be available to cover the decomposer and the substrate with a thin water film. This film provides the transport medium for the organism to invade the substrate, the digestive medium for the extracellular enzymes to work in and the transport medium for the products of decomposition to move to the decomposer and other waste products to leave the site of decomposition (e.g., CO2).
There is a succession of opportunistic decomposer organisms, with new microbial biomass will growing until its supply of substrate is exhausted. As a result of bacterial growth, the population of protozoan graziers will increase. Other predators of the initial decomposers will also increase. Fungi are commonly initial colonisers because their hyphal growth habit allows them to ramify (like a root growing in soil) through the organic substrate whereas bacteria must move where the thin water film allows them to swim. Various types of bacteria and fungi will predate upon each other, each possessing extracellular enzymes or antibiotic agents capable of lysing (breaking down) the cell walls of their prey. The isolation and culture of such organisms on synthetic media has allowed the production of antibiotics (e.g., penicillin and streptomycin) for treatment of human bacterial and fungal infections.
Heterotrophic bacteria
Those which require organic compounds to supply their energy needs are the most common, obtaining these components by secreting externally released extracellular enzymes which attack live and dead animal and plant material. The extracellular enzymes break down the more complex organic materials to simple compounds that can be absorbed by the bacteria.
Fortunately bacteria can attack a wide range of organic substrates, including many organic pesticides and herbicides which are added to crops. Thus toxic substances are prevented from accumulating in the soil.
Bacteria have to compete for inorganic nutrients with plants and fungi and absorb their nitrate, sulphate, phosphate, calcium, potassium, magnesium, and sodium from soil solution.
Nitrification.
A process whereby ammonium N is converted to nitrite by the bacteria Nitrosomonas and then nitrate by the bacteria Nitrobacter. Although they grow slowly, large quantities of ammonium can be converted to nitrate in soil. Rates of nitrification are higher in moist neutral to slightly alkaline soils. Oxygen is required for growth. The nitrate produced can then be taken up by plants, lost by leaching or denitrification (an anaerobic process in water logged soil or in the anaerobic centre of soil aggregates in aerobic conditions.
Nitrogen is an essential element for the growth and functioning of all organisms. Soil parent materials contain little or no nitrogen and the major source of nitrogen is inert N2 gas in the atmosphere. This inert N source must initially be converted into a form which can be used by producer organisms (NH3, NH4+ or NO3-). In the natural world this task falls to a few lightning bolts and specialised micro-organisms capable of fixing molecular N2 into NH3. The rest of the organisms on earth are totally dependent upon these micro-organisms and the few producer organisms capable of catalysing the synthesis of essential amino acids. The importance of organisms capable of fixing N2 gas and knowledge of their maintenance are an essential part of global sustainability. In particular the economies of Australia and New Zealand are heavily dependent upon the successful cultivation of legume based pastures.
It has been estimated that in New Zealand approximately 1 million tonnes of N is fixed annually in legume based pastures. In soils where the concentration of inorganic N is high, clovers will use soil N, with the result that there is less biological fixation of atmospheric N. Nitrogen fertiliser addition to pasture soils has often been shown to decrease BNF (Table 4.5.3). With increasing N addition N fixation by clover continue to decrease and the percent decrease varied between 30 to 70% depending on the time of application and the grazing management.
Nitrogen fixation in soils
Fungi
Fungi are heterotrophs. They are as important as bacteria as decomposers in neutral to alkaline soils but play the dominant decomposing role in the soils of pH <5, which are not suited for bacterial growth. Fungi grow from spores by a threadlike structure called hypha (plural hyphae). A mass of hyphal threads is called a mycelium and is the growing, feeding part of the fungus. Mycelia grow into and invade organic material. In favourable, moist conditions fungi can spread very rapidly e.g. bread mould. Typical hyphae are 5 μm wide and the extreme can be meters in length. Fungi are most predominant in the litter layers of soils and are either saprophytes (feeding on dead organic matter), parasites (feeding on living plants or animals) and symbionts (parasites with beneficial effects for the host). Like bacteria, fungi can use a wide range of organic compounds for food. Fungi are the major decomposers of lignin – the major component of wood and secondary thickening in plants and are therefore mainly responsible for the rotting of wood.
Although many fungi are useful decomposers, others can be harmful, causing economically important root diseases especially to young seedlings. Others are obligate symbionts. Symbiotic associations between fungi and roots are termed mycorrhizae.
Symbiotic associations formed by fungi with roots, and known as mycorrhizae (Gr myces = fungus, rhizo = roots) are of particular importance in the uptake of phosphorus (P). The beneficial effect of mycorrhizae on plant growth has mostly been attributed to an increase in the uptake of nutrients, especially phosphorus (P).
The mycorrhizal association usually increases the growth of plants solely by enhancing the uptake of nutrients, especially that of P. Mycorrhizal fungi in association with plant roots seem likely to increase P uptake by increasing the exploration of soil volume, thereby making 'positionally unavailable' nutrients 'available'. This is achieved by decreasing the distance for diffusion of phosphate ions and by increasing the surface area for absorption.
Although many fungi are useful decomposers, others can be harmful, causing economically important root diseases especially to young seedlings. Others are obligate symbionts. Symbiotic associations between fungi and roots are termed mycorrhizae.
Symbiotic associations formed by fungi with roots, and known as mycorrhizae (Gr myces = fungus, rhizo = roots) are of particular importance in the uptake of phosphorus (P). The beneficial effect of mycorrhizae on plant growth has mostly been attributed to an increase in the uptake of nutrients, especially phosphorus (P).
The mycorrhizal association usually increases the growth of plants solely by enhancing the uptake of nutrients, especially that of P. Mycorrhizal fungi in association with plant roots seem likely to increase P uptake by increasing the exploration of soil volume, thereby making 'positionally unavailable' nutrients 'available'. This is achieved by decreasing the distance for diffusion of phosphate ions and by increasing the surface area for absorption.
Protozoa
These are the smallest of soil animals and prey on soil bacteria and to some extent fungi. Protozoa are heterotrophic, unicellular, motile, usually non-photosynthetic organisms. They generally feed on dead bacterial cells and organic matter. Some ingest solid food by engulfing (phagocytosis). Some eat live bacteria. Few use only water-soluble compounds.
Protozoa reproduce both asexually (by binary fission) and sexually. They are ecologically important in organic matter decomposition.
Protozoa reproduce both asexually (by binary fission) and sexually. They are ecologically important in organic matter decomposition.
Arthropods
This group includes mites, springtails, beetles, many insect larvae, ant, termites, millipedes and centipedes.
Mites and springtails are often the most numerous feeding on detritus in the lower litter of the forests and undisturbed grassland. Some species of mite are predatory on springtails and nematodes. The most important role is that of fragmenting litter, leading to easier decay by fungi and bacteria.
Many beetles and insect larvae live in soils and are serious pests of agricultural crops. Some beetles, however, are beneficial in that they mix animal dung into the soil. One species of dung beetle has been introduced into Northland to improve the return of sheep and cattle dung to the land.
Millipedes feed on vegetation and aid the return of organic material to the soil, whereas centipedes are carnivorous and return soil animal material to the soil via their faeces.
Arthropods do not mix plant and animal material intimately with soil minerals as they do not ingest mineral material.
Mites and springtails are often the most numerous feeding on detritus in the lower litter of the forests and undisturbed grassland. Some species of mite are predatory on springtails and nematodes. The most important role is that of fragmenting litter, leading to easier decay by fungi and bacteria.
Many beetles and insect larvae live in soils and are serious pests of agricultural crops. Some beetles, however, are beneficial in that they mix animal dung into the soil. One species of dung beetle has been introduced into Northland to improve the return of sheep and cattle dung to the land.
Millipedes feed on vegetation and aid the return of organic material to the soil, whereas centipedes are carnivorous and return soil animal material to the soil via their faeces.
Arthropods do not mix plant and animal material intimately with soil minerals as they do not ingest mineral material.
Earthworms
Earthworms feed exclusively on dead organic matter which they ingest with large quantities of clay and silt size particles. As a result, in soils with large earthworm populations, organic and mineral matter are more homogeneously mixed to a greater depth than if the earthworms are absent.
There are over 190 earthworm species in New Zealand of which only a few introduced species (Aporrectodea caliginosa and Lumbricus rubellus) are considered to be beneficial. The number of earthworm population fluctuates with season which is related to the soil moisture, temperature and supply of food.
The most agronomically beneficial earthworms in New Zealand are European imports of the Lumbricidae family. These earthworms are not as large as some native bush worms (of the Megascolecidae family) but are more active in incorporating plant and animal excreta into topsoils. They spend most of their time in moist topsoils (>60% field capacity) but in order to survive will retreat to an inactive state in the subsoil as the topsoil dries out. Thus earthworm activity is restricted to periods of high rainfall and cooler temperatures (optimum about 12°C) as evidenced by peak casting activity in May-June and September-October.
In neutral soils rich in exchangeable calcium there can be as many as 7*10-6 per hectare (2400 kg/ha) which is a live weight equivalent approximately 40 ewes/ha. When populations are this large and include surface casting species such as Aporrectodea, up to 40 tonnes of cast material can be deposited on the soil surface annually. If soil pH falls below 4.5, earthworms are often absent.
Earthworms promote the aggregate stability of soil either through the production of gums (polysaccharides) and/or through the binding effect of fungal hyphae in the casts.
Earthworm affects the pore space in soils by burrowing and by depositing their casts as loosely packed aggregates on the soil surface. Up to two-thirds of the air-filled pore space in soils may be earthworm burrows. These burrows are known as “macropores” which increases the rate of infiltration.
Earthworms play a major role in the removal from surface of plant litter, dung and other organic material and its incorporation into soils.
There are over 190 earthworm species in New Zealand of which only a few introduced species (Aporrectodea caliginosa and Lumbricus rubellus) are considered to be beneficial. The number of earthworm population fluctuates with season which is related to the soil moisture, temperature and supply of food.
The most agronomically beneficial earthworms in New Zealand are European imports of the Lumbricidae family. These earthworms are not as large as some native bush worms (of the Megascolecidae family) but are more active in incorporating plant and animal excreta into topsoils. They spend most of their time in moist topsoils (>60% field capacity) but in order to survive will retreat to an inactive state in the subsoil as the topsoil dries out. Thus earthworm activity is restricted to periods of high rainfall and cooler temperatures (optimum about 12°C) as evidenced by peak casting activity in May-June and September-October.
In neutral soils rich in exchangeable calcium there can be as many as 7*10-6 per hectare (2400 kg/ha) which is a live weight equivalent approximately 40 ewes/ha. When populations are this large and include surface casting species such as Aporrectodea, up to 40 tonnes of cast material can be deposited on the soil surface annually. If soil pH falls below 4.5, earthworms are often absent.
Earthworms promote the aggregate stability of soil either through the production of gums (polysaccharides) and/or through the binding effect of fungal hyphae in the casts.
Earthworm affects the pore space in soils by burrowing and by depositing their casts as loosely packed aggregates on the soil surface. Up to two-thirds of the air-filled pore space in soils may be earthworm burrows. These burrows are known as “macropores” which increases the rate of infiltration.
Earthworms play a major role in the removal from surface of plant litter, dung and other organic material and its incorporation into soils.
Nematodes
The soil contains enormous numbers of nematodes. They are usually inactive unless the soil has free water. They range from free living organic matter feeders, to highly specialised parasites of plants, animals and even man. The structure of the mouth or buccal capsule will indicate their mode of life.
Nematodes are pests (pathogenic) of many horticultural crops and can be a nuisance in bulb production and root crops. In this respect, parasitism of plants by nematodes may be the first step in the death of the plant and the decay process. Hence although nematodes may be seen to speed up the return of the nutrients to the soil, their major economic importance is that of a plant pest.
Nematodes are pests (pathogenic) of many horticultural crops and can be a nuisance in bulb production and root crops. In this respect, parasitism of plants by nematodes may be the first step in the death of the plant and the decay process. Hence although nematodes may be seen to speed up the return of the nutrients to the soil, their major economic importance is that of a plant pest.
Extra Readings
|
| ||||